1. Disease Information
1.1 Disease overview (what it is)
The disorder is described as severe microlissencephaly due to biallelic pathogenic variants in KATNB1, encoding the p80 regulatory subunit of the katanin microtubule-severing complex. Affected individuals show severe microcephaly with simplified cortical gyri/sulci, typically with dramatically reduced cortical volume on MRI. (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates pages 1-2)
1.2 Key identifiers and controlled names
- Gene: KATNB1; a microcephaly-disorders review explicitly notes KATNB1 (MIM# 602703) and uses the disease label “Lissencephaly 6 with microcephaly (LIS6)”. (brown2017geneticrequirementsfor pages 103-108)
- Standardized disease name used in primary literature: “lissencephaly 6” and “microlissencephaly”. (peluso2021wholeexomesequencing pages 1-2, hu2014kataninp80regulates pages 2-4)
- MONDO / Orphanet / ICD / MeSH / OMIM disease-number: Not present in the retrieved full-text excerpts; therefore not reported here to avoid mis-mapping. (peluso2021wholeexomesequencing pages 1-2, brown2017geneticrequirementsfor pages 103-108)
1.3 Synonyms and alternative names
- KATNB1-related microlissencephaly (primary literature framing) (hu2014kataninp80regulates pages 2-4)
- Lissencephaly 6 with microcephaly (LIS6) (brown2017geneticrequirementsfor pages 103-108, peluso2021wholeexomesequencing pages 1-2)
- KATNB1-related cortical malformation (umbrella term consistent with MCD phenotype) (peluso2021wholeexomesequencing pages 6-8)
1.4 Evidence sources (individual vs aggregated)
The knowledge base here is derived from: - Primary human family-based studies describing affected individuals and segregating variants (Hu et al., Neuron 2014). (hu2014kataninp80regulates pages 2-4) - Case report plus literature aggregation with pooled counts across published patients (Peluso et al., Genes 2021). (peluso2021wholeexomesequencing pages 6-8) - Mechanistic studies using patient-derived iPSCs/brain organoids and animal models (Jin et al., 2017; Hu et al., 2014). (jin2017kataninp80numa pages 1-3, hu2014kataninp80regulates pages 1-2) - Consensus diagnostic guidance for MCDs (Neuro-MIG; Oegema et al., Nat Rev Neurol 2020). (oegema2020internationalconsensusrecommendations pages 8-9, oegema2020internationalconsensusrecommendations pages 7-8)
2. Etiology
2.1 Disease causal factors
Primary cause: Biallelic pathogenic variants in KATNB1 disrupting normal katanin p80 function, leading to abnormal cortical development. (hu2014kataninp80regulates pages 2-4, peluso2021wholeexomesequencing pages 6-8)
Inheritance: Evidence supports autosomal recessive inheritance, including consanguineous pedigrees and parental heterozygosity for proband homozygous variants. (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 1-2)
2.2 Risk factors
- Genetic risk factor: Having biallelic (homozygous or compound heterozygous) loss-of-function KATNB1 variants. (peluso2021wholeexomesequencing pages 6-8)
- Family structure: Consanguinity increases risk for recessive disorders; KATNB1 cases include first-cousin unions in reported families. (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 1-2)
Environmental risk factors and infectious triggers were not identified as causal for this monogenic condition in the retrieved evidence.
2.3 Protective factors
No genetic or environmental protective factors have been reported in the retrieved evidence.
2.4 Gene–environment interactions
No KATNB1-specific gene–environment interaction evidence was found in the retrieved corpus.
3. Phenotypes (clinical features)
3.1 Core phenotype spectrum
Across three unrelated families, affected individuals had severe microcephaly, global developmental delay, and seizures, with MRI showing markedly reduced cortical volume and simplified gyral pattern. (hu2014kataninp80regulates pages 1-2)
A literature aggregation summarized 13 subjects from nine families (age range 11 months–12 years) and emphasized microcephaly as characteristic. (peluso2021wholeexomesequencing pages 6-8)
3.2 Quantitative outcome and frequency data (from available aggregation)
From the Peluso 2021 aggregation: - Head circumference at birth: approximately −1.63 to −5.9 SD, with further decline in many patients. (peluso2021wholeexomesequencing pages 6-8) - Progression: in 8/14 cases head circumference declined to < −6 SD (and one reported < −11 SD by 12 years). (peluso2021wholeexomesequencing pages 6-8) - Neurodevelopmental outcomes: - Regular/normal psychomotor development reported in 2 patients. - 5 patients never achieved independent walking. - 5 patients had absent speech. (peluso2021wholeexomesequencing pages 6-8) - Hypertonia: reported in 8/14, mainly lower limbs. (peluso2021wholeexomesequencing pages 6-8)
Seizure frequency (percent affected) was not quantified in the retrieved aggregation excerpts.
3.3 Neuroimaging phenotypes
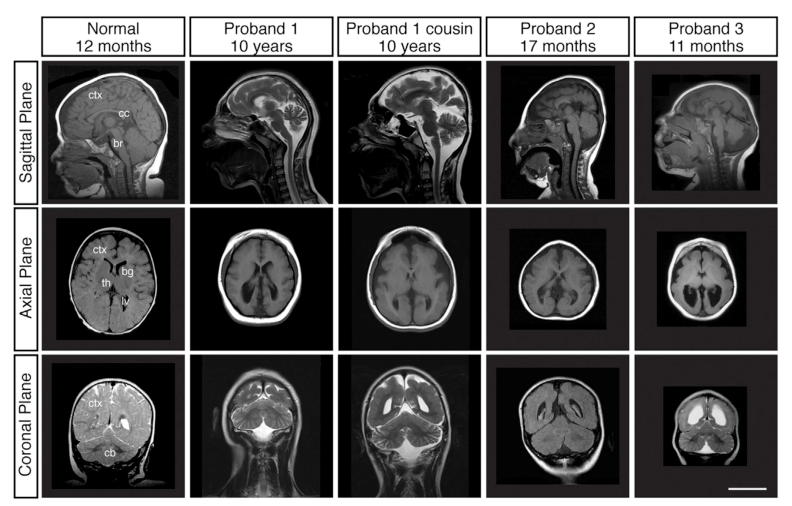
MRI findings in primary and aggregated reports include: - Reduced cortical size/volume and simplified gyral folding with shallow sulci. (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates media 9e221719) - Posterior ventriculomegaly (enlarged lateral ventricles posteriorly). (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates media 9e221719) - Corpus callosum thinning/abnormalities. (hu2014kataninp80regulates pages 2-4, peluso2021wholeexomesequencing pages 6-8) - Additional reported cortical malformations across cases: pachygyria, polymicrogyria, and periventricular/bilateral nodular heterotopia. (peluso2021wholeexomesequencing pages 6-8)
3.4 Suggested HPO terms (examples)
Key HPO mappings are provided in the ontology table artifact. (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 6-8)
3.5 Quality-of-life impact
Formal QoL instruments (EQ-5D/SF-36/PROMIS) were not reported in the retrieved evidence. Functional impact is inferred from high rates of absent speech/non-walking and severe microcephaly. (peluso2021wholeexomesequencing pages 6-8)
4. Genetic/Molecular Information
4.1 Causal gene
- KATNB1 encodes the p80 regulatory subunit of katanin, which regulates the localization/activity of the catalytic p60 subunit in a heterodimeric microtubule-severing complex. (lynn2021themammalianfamily pages 1-2, lynn2023methodsandsystems pages 20-26)
4.2 Pathogenic variant types and examples
Primary human study (Hu et al., 2014) identified homozygous deleterious variants across three families: - Start-codon (initiator ATG)–abolishing variant. (hu2014kataninp80regulates pages 2-4) - Conserved glycine-to-tryptophan missense variant in WD40 region. (hu2014kataninp80regulates pages 2-4) - Splice donor variant at exon 6 boundary causing exon 6 skipping (dele6). (hu2014kataninp80regulates pages 2-4)
Case report (Peluso et al., 2021): - Homozygous splice-site variant NM_005886.3:c.1416+1del (absent from gnomAD v2.1.1 at time of report), with parental heterozygosity. (peluso2021wholeexomesequencing pages 4-6, peluso2021wholeexomesequencing pages 1-2)
Additional variants mentioned in a microcephaly disorders review (secondary synthesis; should be traced to primary sources for clinical interpretation): S535L, L540R, V45I, V150Cfs*22. (brown2017geneticrequirementsfor pages 103-108)
4.3 Functional consequences
Evidence supports predominantly loss-of-function/hypomorphic mechanisms: - Hu et al. report reduced protein levels and functional defects (e.g., dele6 stable but functionally defective; altered localization). (hu2014kataninp80regulates pages 2-4) - KATNB1 is under strong negative selection (Ka/Ks ~0.03–0.08), consistent with functional constraint. (hu2014kataninp80regulates pages 2-4)
4.4 Modifier genes / epigenetics / chromosomal abnormalities
No validated KATNB1-specific modifier genes, epigenetic signatures, or recurrent chromosomal abnormalities were identified in the retrieved evidence.
5. Environmental Information
No established environmental, lifestyle, or infectious causal contributors to KATNB1-related microlissencephaly were identified in the retrieved evidence.
6. Mechanism / Pathophysiology
6.1 Key concepts (current understanding)
KATNB1 (p80) is part of the katanin microtubule-severing system. In cortical development, current evidence links KATNB1 dysfunction to centrosome/centriole abnormalities, aberrant ciliogenesis, mitotic spindle defects, and downstream disruption of morphogen signaling—ultimately impairing neural progenitor divisions and neuronal migration. (hu2014kataninp80regulates pages 1-2, jin2017kataninp80numa pages 1-3, lynn2023methodsandsystemsa pages 31-36)
6.2 Causal chain (from variant to phenotype)
- Biallelic KATNB1 loss-of-function → impaired regulation/localization of katanin activity and microtubule remodeling. (lynn2021themammalianfamily pages 1-2, lynn2023methodsandsystems pages 20-26)
- Centrosome/centriole dysregulation → supernumerary centrioles, multipolar spindles, and ectopic/supernumerary primary cilia. (hu2014kataninp80regulates pages 2-4)
- Cilia dysfunction → impaired Hedgehog signaling (e.g., reduced GLI1/Patched expression reported in summaries of KATNB1-null contexts). (lynn2023methodsandsystems pages 41-46, hu2014kataninp80regulates pages 1-2)
- Neural stem/progenitor defects (reduced proliferation, increased cell death, altered asymmetric division) → reduced cortical mass and microcephaly. (zaidi2022primaryciliainfluence pages 9-10, hu2014kataninp80regulates pages 1-2)
- Neuronal migration defects (and disturbed neurogenesis) → simplified gyral pattern/lissencephaly-spectrum malformation and associated epilepsy/developmental disability. (jin2017kataninp80numa pages 1-3)
6.3 Cellular processes and pathways
- Mitotic microtubule/spindle organization: KATNB1 is essential for aster formation/maintenance; depletion produces abnormal mitoses and neurodevelopmental abnormalities. (jin2017kataninp80numa pages 1-3)
- Centrosome and cilium biology: KATNB1 regulates centriole (including mother centriole) and cilia number; loss leads to excess centrioles and cilia. (hu2014kataninp80regulates pages 1-2)
- Hedgehog signaling: Perturbed Shh/Hedgehog signaling accompanies KATNB1 loss. (hu2014kataninp80regulates pages 2-4)
6.4 Evidence from advanced technologies
Patient-derived iPSCs and brain organoids recapitulate mitotic/microtubule and neuronal migration defects observed in experimental depletion models, supporting human relevance. (jin2017kataninp80numa pages 1-3)
6.5 Suggested GO and CL terms
A structured list is provided in the ontology artifact. (hu2014kataninp80regulates pages 1-2, duy2024the"microcephalichydrocephalus" pages 6-7)
7. Anatomical Structures Affected
7.1 Organ/system level
- Central nervous system, especially cerebral cortex (reduced cortical volume, simplified gyration). (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates media 9e221719)
7.2 Tissue/cell level
Neural stem/progenitor compartments implicated include neuroepithelial cells and radial glia (NSC paradigm emphasized in recent synthesis; cortical progenitor/cilium context described in primary paper). (duy2024the"microcephalichydrocephalus" pages 6-7, hu2014kataninp80regulates pages 2-4)
7.3 Subcellular level
- Centrosome/centriole and primary cilium, and mitotic spindle are central subcellular structures implicated by functional studies. (hu2014kataninp80regulates pages 1-2, lynn2023methodsandsystemsa pages 31-36)
8. Temporal Development
8.1 Onset
Evidence is consistent with congenital onset, with microcephaly present at birth in reported cases and MRI demonstrating early developmental malformation. (peluso2021wholeexomesequencing pages 6-8, hu2014kataninp80regulates pages 1-2)
8.2 Progression
Microcephaly can be progressive postnatally in many individuals (declining SD over time), while neurodevelopmental impairments can be severe and persistent. (peluso2021wholeexomesequencing pages 6-8)
9. Inheritance and Population
9.1 Inheritance pattern
Autosomal recessive inheritance is supported by consanguinity, homozygous variants, and parental carrier status. (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 1-2)
9.2 Epidemiology
No prevalence/incidence estimates were identified in the retrieved evidence.
9.3 Population genetics and rarity indicators
- The c.1416+1del splice variant was absent from gnomAD v2.1.1 at time of reporting, supporting ultra-rarity. (peluso2021wholeexomesequencing pages 4-6)
- KATNB1 shows strong evolutionary constraint (Ka/Ks ~0.03–0.08), suggesting intolerance to nonsynonymous variation. (hu2014kataninp80regulates pages 2-4)
10. Diagnostics
10.1 Clinical diagnosis
Diagnosis is suggested by congenital microcephaly with MRI evidence of microlissencephaly/simplified gyral pattern and associated MCD features such as ventriculomegaly and corpus callosum thinning. (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates media 9e221719)
10.2 Genetic testing approach (expert guidance)
Neuro-MIG international consensus recommends that all individuals with MCD pursue an etiologic diagnosis using: - Expert MRI review and pattern recognition to guide testing. - Chromosomal microarray (CMA) as first-tier test. - NGS panels or trio exome/genome to maximize yield; consider deep sequencing for mosaicism, segregation testing, multi-tissue testing, and functional validation when needed. (oegema2020internationalconsensusrecommendations pages 7-8, oegema2020internationalconsensusrecommendations pages 12-13)
10.3 KATNB1-specific implementation example
A targeted NGS panel for brain malformations/microcephaly detected a homozygous KATNB1 splice variant, with confirmatory Sanger sequencing, segregation in parents, and checks against public databases (gnomAD/HGMD). (peluso2021wholeexomesequencing pages 4-6)
11. Outcome/Prognosis
Outcome is variable but often severe: - Among aggregated cases: severe motor and speech impairment is common (non-walking and absent speech each reported in 5 patients), though normal psychomotor development has been reported in 2 patients. (peluso2021wholeexomesequencing pages 6-8) - Hypertonia is frequent (8/14). (peluso2021wholeexomesequencing pages 6-8)
Formal survival/life expectancy data were not available in the retrieved evidence.
12. Treatment
12.1 Disease-modifying therapies
No disease-modifying or gene-targeted therapies were identified in the retrieved evidence for KATNB1-related microlissencephaly.
12.2 Current clinical management (real-world implementation)
Evidence supports supportive, multidisciplinary care, including: - Neurologic care for seizures (seizures are a common feature, but specific antiseizure medication regimens were not described in available excerpts). (hu2014kataninp80regulates pages 1-2) - MRI-based monitoring/characterization and multi-system evaluations (ophthalmologic, audiologic, cardiologic) as seen in reported cases. (peluso2021wholeexomesequencing pages 4-6) - Supportive interventions for complications (e.g., albumin infusions for nephrotic syndrome in one reported case, although this complication is not established as core/typical in all KATNB1 cases). (peluso2021wholeexomesequencing pages 4-6)
12.3 Suggested MAXO terms
Suggested MAXO terms are included in the ontology artifact; these represent standard care actions inferred from phenotype and diagnostic practice. (oegema2020internationalconsensusrecommendations pages 7-8, peluso2021wholeexomesequencing pages 6-8)
13. Prevention
No primary prevention is available for this genetic disorder. Prevention in practice is genetic counseling and carrier/recurrence-risk assessment due to autosomal recessive inheritance and observed consanguinity in multiple families. (peluso2021wholeexomesequencing pages 1-2, hu2014kataninp80regulates pages 1-2)
14. Other Species / Natural Disease
No naturally occurring veterinary analogs were identified in the retrieved evidence.
15. Model Organisms
Evidence supports multiple model systems: - Mouse: Katnb1 loss is associated with essential roles in neurogenesis and cell survival and Shh-related developmental abnormalities (e.g., holoprosencephaly hallmarks described in the primary report’s summary). (hu2014kataninp80regulates pages 1-2) - Zebrafish: loss of zebrafish katnb1 reveals roles in early and late developmental stages (reported in the primary study summary). (hu2014kataninp80regulates pages 1-2) - Human iPSC/brain organoids: patient-derived KATNB1-mutant iPSCs and brain organoids confirm mitotic and neuronal migration defects. (jin2017kataninp80numa pages 1-3)
Figures and structured summaries
The following artifact summarizes the best-supported clinical, genetic, imaging, and mechanistic facts, including key case counts and variant examples.
Table (click to expand)
| Aspect | Key details (concise) | Evidence/source (include PMID/DOI and year) |
|---|---|---|
| Disease framing / synonyms | KATNB1-related cortical malformation is described as severe congenital microlissencephaly / lissencephaly 6 with combined microcephaly and lissencephaly-spectrum cortical malformation. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Peluso et al., Genes (2021), DOI: 10.3390/genes12070962; Jin et al., Sci Rep (2017), DOI: 10.1038/srep39902 (hu2014kataninp80regulates pages 2-4, peluso2021wholeexomesequencing pages 1-2, jin2017kataninp80numa pages 1-3) |
| Inheritance | Reported as autosomal recessive; families include consanguineous pedigrees and affected individuals with homozygous or compound heterozygous variants; parental heterozygosity shown for c.1416+1del. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Peluso et al., Genes (2021), DOI: 10.3390/genes12070962 (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 1-2, peluso2021wholeexomesequencing pages 4-6) |
| Core clinical features | Severe congenital microcephaly, global developmental delay / psychomotor impairment, seizures, hypertonia (especially lower limbs in aggregated series), and variable absent speech / failure to achieve independent walking. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Peluso et al., Genes (2021), DOI: 10.3390/genes12070962 (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 6-8) |
| Head-size severity | Microcephaly is characteristic; aggregated summary reported head circumference from about -1.63 to -5.9 SD at birth, with further decline in many patients; 8/14 reportedly fell below -6 SD. | Peluso et al., Genes (2021), DOI: 10.3390/genes12070962 (peluso2021wholeexomesequencing pages 6-8) |
| Neuroimaging pattern | Reduced cortical size / brain volume with simplified gyral pattern, shallow sulci, posteriorly enlarged lateral ventricles, thinning or abnormalities of the corpus callosum; relative sparing of cerebellum, basal ganglia, thalamus, and brainstem in the original Neuron report. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017 (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates pages 1-2) |
| Additional imaging findings across reports | Pachygyria, polymicrogyria, simplified gyral pattern, periventricular or bilateral nodular heterotopia, posterior fossa anomalies; in one 2021 case, subarachnoid dilatation, abnormal gyration, slight hippocampal malrotation, subcortical heterotopia, and slightly thickened cortex. | Peluso et al., Genes (2021), DOI: 10.3390/genes12070962 (peluso2021wholeexomesequencing pages 6-8, peluso2021wholeexomesequencing pages 4-6) |
| Reported case counts | Hu et al. studied 3 families / 3 probands in detail; one family reportedly had 5 affected individuals. Peluso summarized prior literature as 13 subjects from 9 families, while some aggregated phenotype counts were tabulated over 14 cases (including the newly reported case). | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Peluso et al., Genes (2021), DOI: 10.3390/genes12070962 (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 6-8) |
| Example pathogenic variants from primary reports | Homozygous start-codon–abolishing variant; homozygous missense variant in a conserved WD40 repeat (glycine to tryptophan); homozygous splice-donor variant causing exon 6 skipping (dele6); homozygous splice-site variant NM_005886.3:c.1416+1del. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Peluso et al., Genes (2021), DOI: 10.3390/genes12070962 (hu2014kataninp80regulates pages 2-4, peluso2021wholeexomesequencing pages 4-6) |
| Other reported variant examples in literature summary | Additional literature summary lists homozygous pathogenic variants including S535L, L540R, V45I, and frameshift V150Cfs*22, supporting loss-of-function / severe functional impairment. | Brown review summary citing Mishra-Gorur/Hu era literature (2017) (brown2017geneticrequirementsfor pages 103-108) |
| Population rarity | The 2021 splice variant c.1416+1del was absent from gnomAD v2.1.1 and HGMD at time of report; the original 2014 variants were absent from matched control populations. | Peluso et al., Genes (2021), DOI: 10.3390/genes12070962; Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017 (peluso2021wholeexomesequencing pages 4-6, hu2014kataninp80regulates pages 2-4) |
| KATNB1 molecular role | KATNB1 encodes the p80 regulatory B-subunit of the katanin microtubule-severing complex; it regulates localization/activity of the catalytic A-subunit and is important for corticogenesis. | Lynn et al., Front Cell Dev Biol (2021), DOI: 10.3389/fcell.2021.692040 (lynn2021themammalianfamily pages 1-2, lynn2021themammalianfamily pages 14-15, lynn2021themammalianfamily pages 8-10) |
| Mechanistic theme: centriole / cilia control | KATNB1 loss causes excess centrioles, increased mother centrioles, supernumerary cilia / aberrant ciliogenesis, linking disease to centrosome-cilia homeostasis defects. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Zaidi et al., Cells (2022), DOI: 10.3390/cells11182895 (hu2014kataninp80regulates pages 1-2, zaidi2022primaryciliainfluence pages 9-10) |
| Mechanistic theme: spindle / mitotic defects | Patient-derived or depleted cells show defective proliferation, abnormal mitotic spindles, aster-formation defects, reduced spindle-pole microtubules, supernumerary centrosomes, and cytokinesis-related abnormalities. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Jin et al., Sci Rep (2017), DOI: 10.1038/srep39902; Lynn methods summary (2023) (hu2014kataninp80regulates pages 1-2, jin2017kataninp80numa pages 1-3, lynn2023methodsandsystemsa pages 31-36) |
| Mechanistic theme: Hedgehog signaling | Katnb1-null cells show defective Hedgehog signaling with reduced GLI1 and Patched expression, supporting a cilia-dependent developmental signaling defect upstream of impaired corticogenesis. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Lynn methods summary (2023) (hu2014kataninp80regulates pages 1-2, lynn2023methodsandsystems pages 41-46) |
| Mechanistic theme: neurogenesis / migration | Loss or depletion impairs neurogenesis, reduces neural progenitor proliferation, increases cell death in some models, and disrupts neuronal migration; disease pathogenesis is linked to abnormal asymmetrically dividing neural progenitors. | Hu et al., Neuron (2014), DOI: 10.1016/j.neuron.2014.12.017; Jin et al., Sci Rep (2017), DOI: 10.1038/srep39902; Zaidi et al., Cells (2022), DOI: 10.3390/cells11182895 (hu2014kataninp80regulates pages 1-2, jin2017kataninp80numa pages 1-3, zaidi2022primaryciliainfluence pages 9-10) |
| Human stem-cell / organoid evidence | Findings were confirmed in patient-derived KATNB1-mutant iPSCs and brain organoids, supporting relevance of spindle/MT and migration defects to human cortical development. | Jin et al., Sci Rep (2017), DOI: 10.1038/srep39902 (jin2017kataninp80numa pages 1-3) |
Table: This table concisely summarizes the clinical, genetic, imaging, and mechanistic evidence for KATNB1-related cortical malformation using only the provided evidence snippets. It is useful as a structured reference for a disease knowledge base entry on microlissencephaly / lissencephaly 6.
Ontology term suggestions (HPO/GO/CL/UBERON/MAXO) are summarized here:
Table (click to expand)
| Category | Suggested term label | Suggested ID | Rationale/notes linked to evidence |
|---|---|---|---|
| HPO | Microcephaly | HP:0000252 | Core feature across reported families/cases; severe congenital microcephaly is repeatedly emphasized in KATNB1-related microlissencephaly/lissencephaly 6 (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 6-8). |
| HPO | Seizure | HP:0001250 | Seizures are reported among major presenting neurological features in affected individuals (hu2014kataninp80regulates pages 1-2). |
| HPO | Global developmental delay | HP:0001263 | Human cases show global developmental delay / severe psychomotor impairment, with delayed walking and speech or absent milestones in several patients (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 6-8). |
| HPO | Lissencephaly | HP:0001339 | Disease is framed as microlissencephaly / lissencephaly 6; simplified cortical folding is central to diagnosis (hu2014kataninp80regulates pages 2-4, peluso2021wholeexomesequencing pages 1-2). |
| HPO | Simplified gyral pattern | HP:0009879 | MRI in affected individuals shows simplification of gyral folding pattern with shallow sulci (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates media 9e221719). |
| HPO | Ventriculomegaly | HP:0002119 | Enlarged lateral ventricles, particularly posteriorly, are described on MRI (hu2014kataninp80regulates pages 2-4). |
| HPO | Abnormality of the corpus callosum | HP:0001273 | Corpus callosum thinning/abnormality is repeatedly reported in neuroimaging summaries (peluso2021wholeexomesequencing pages 6-8, hu2014kataninp80regulates pages 2-4). |
| HPO | Hypertonia | HP:0001276 | Hypertonia, particularly affecting lower limbs, was reported in aggregated clinical summaries (peluso2021wholeexomesequencing pages 6-8). |
| HPO | Periventricular nodular heterotopia | HP:0002136 | Periventricular/bilateral nodular heterotopia has been reported among associated cortical malformations (peluso2021wholeexomesequencing pages 6-8, peluso2021wholeexomesequencing pages 4-6). |
| GO | microtubule severing | GO:0051013 | KATNB1 encodes the regulatory p80 subunit of katanin, a microtubule-severing complex; this is central to current molecular understanding (lynn2021themammalianfamily pages 1-2, lynn2023methodsandsystems pages 20-26). |
| GO | mitotic spindle organization | GO:0007052 | KATNB1 loss causes spindle defects, abnormal mitoses, reduced spindle-pole microtubules, and aster defects (hu2014kataninp80regulates pages 1-2, jin2017kataninp80numa pages 1-3, lynn2023methodsandsystemsa pages 31-36). |
| GO | centriole duplication | GO:0031534 | Katnb1-null or depleted cells show excess centrioles / centriole overduplication, directly supporting this process as disease-relevant (hu2014kataninp80regulates pages 1-2, lynn2023methodsandsystems pages 41-46). |
| GO | cilium assembly | GO:0060271 | Supernumerary cilia / aberrant ciliogenesis are a recurring mechanistic finding in KATNB1-deficient systems (hu2014kataninp80regulates pages 1-2, lynn2023methodsandsystems pages 41-46, lynn2023methodsandsystemsa pages 41-46). |
| GO | Hedgehog signaling pathway | GO:0007224 | Defective Hedgehog signaling with reduced GLI1/Patched expression is reported in KATNB1-deficient models (hu2014kataninp80regulates pages 1-2, lynn2023methodsandsystems pages 41-46). |
| GO | neuron migration | GO:0001764 | Patient-derived iPSC/organoid and mouse studies show impaired neuronal migration after KATNB1 disruption (jin2017kataninp80numa pages 1-3, lynn2023methodsandsystemsa pages 41-46). |
| GO | neurogenesis | GO:0022008 | Loss of KATNB1 impairs neurogenesis, progenitor proliferation, and neuronal output during cortical development (hu2014kataninp80regulates pages 1-2, zaidi2022primaryciliainfluence pages 9-10, jin2017kataninp80numa pages 1-3). |
| CL | radial glial cell | CL:0010012 | Reviews and mechanistic syntheses place disease biology in cortical radial glia / apical progenitors and neural stem-cell compartments (duy2024the"microcephalichydrocephalus" pages 6-7, duy2024the"microcephalichydrocephalus" pages 4-5). |
| CL | neuroepithelial cell | CL:0002319 | Early cortical neuroepithelial cells are implicated in the developmental context of KATNB1-related disease and cilium/centriole asymmetry (hu2014kataninp80regulates pages 2-4, duy2024the"microcephalichydrocephalus" pages 4-5). |
| CL | neural progenitor cell | CL:0011020 | Reduced cycling/proliferation of neural progenitors is a key mechanism across human/model evidence (zaidi2022primaryciliainfluence pages 9-10, duy2024the"microcephalichydrocephalus" pages 6-7). |
| CL | neuron | CL:0000540 | Reduced cortical neurons and impaired neuronal migration are downstream disease mechanisms (jin2017kataninp80numa pages 1-3, lynn2023methodsandsystemsa pages 41-46). |
| UBERON | cerebral cortex | UBERON:0000956 | Primary malformed structure; imaging shows markedly reduced cortical size and simplified gyration (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates media 9e221719). |
| UBERON | lateral ventricle | UBERON:0002081 | Lateral ventricular enlargement is a consistent imaging feature (hu2014kataninp80regulates pages 2-4, hu2014kataninp80regulates media 9e221719). |
| UBERON | corpus callosum | UBERON:0000924 | Corpus callosum thinning/abnormality is repeatedly noted on MRI (peluso2021wholeexomesequencing pages 6-8, hu2014kataninp80regulates pages 2-4). |
| UBERON | primary cilium | Evidence strongly supports abnormal cilia number/ciliogenesis, but a stable UBERON ID is not confidently assigned here from available context; include as an anatomical target structure (hu2014kataninp80regulates pages 1-2, lynn2023methodsandsystems pages 41-46). | |
| GO | centrosome | GO:0005813 | KATNB1 localizes to centrosomes and disease mechanisms include centrosome numerical/structural abnormalities; GO cellular component is more appropriate than UBERON for this subcellular structure (lynn2023methodsandsystemsa pages 31-36, brown2017geneticrequirementsfor pages 103-108). |
| MAXO | Antiseizure medication therapy | No KATNB1-specific drug regimen is established, but seizure management is a logical supportive action given recurrent epilepsy/seizures in affected individuals (hu2014kataninp80regulates pages 1-2, peluso2021wholeexomesequencing pages 6-8). | |
| MAXO | Brain magnetic resonance imaging | MRI is central for identifying simplified gyral pattern, ventriculomegaly, and corpus callosum abnormalities in diagnosis/workup (hu2014kataninp80regulates pages 2-4, oegema2020internationalconsensusrecommendations pages 1-2). | |
| MAXO | Exome sequencing | WES is recommended/used for diagnosis in malformations of cortical development and specifically highlighted in consanguineous families (peluso2021wholeexomesequencing pages 4-6, oegema2020internationalconsensusrecommendations pages 7-8). | |
| MAXO | Genetic counseling | Autosomal recessive inheritance with parental heterozygosity supports counseling on recurrence risk and family planning (peluso2021wholeexomesequencing pages 1-2, oegema2020internationalconsensusrecommendations pages 1-2). | |
| MAXO | Physical therapy | Supportive rehabilitation is reasonable for severe motor impairment/hypertonia, although no disease-specific trials were identified in the retrieved evidence (peluso2021wholeexomesequencing pages 6-8). | |
| MAXO | Speech therapy | Many patients have absent or severely delayed speech; supportive speech/language intervention is therefore relevant, though not specifically studied in KATNB1 cohorts (peluso2021wholeexomesequencing pages 6-8). |
Table: This table proposes ontology mappings for phenotypes, mechanisms, cell types, anatomy, and care actions relevant to KATNB1-related cortical malformation. It is designed to support structured knowledge-base annotation while keeping suggestions tied to the available evidence.
An example MRI figure demonstrating reduced cortical size, simplified gyral pattern, ventriculomegaly, and corpus callosum thinning is available from Hu et al. 2014 (Figure 1A). (hu2014kataninp80regulates media 9e221719)
Limitations of this report (evidence gaps)
- Formal disease identifiers (MONDO/Orphanet/ICD/MeSH and OMIM disease-number) were not present in retrieved full-text excerpts; they require database lookups not available in this tool context.
- The major Neuron case series Mishra-Gorur et al. 2015 was not available in full text in the retrieved corpus beyond limited metadata, restricting completeness of variant cataloging and phenotype frequencies.
- No controlled clinical guidelines specific to KATNB1, no clinical trials, and limited treatment details were present in the accessible literature excerpts.
Source URLs and publication dates (from retrieved corpus)
- Hu et al. Neuron — published 2014-12-17; https://doi.org/10.1016/j.neuron.2014.12.017 (hu2014kataninp80regulates pages 1-2)
- Jin et al. Scientific Reports — published 2017-01; https://doi.org/10.1038/srep39902 (jin2017kataninp80numa pages 1-3)
- Oegema et al. Nature Reviews Neurology — published 2020-09; https://doi.org/10.1038/s41582-020-0395-6 (oegema2020internationalconsensusrecommendations pages 8-9)
- Peluso et al. Genes — published 2021-06-24; https://doi.org/10.3390/genes12070962 (peluso2021wholeexomesequencing pages 1-2)
- Duy et al. Cerebral Cortex — published 2024-11; https://doi.org/10.1093/cercor/bhad432 (duy2024the"microcephalichydrocephalus" pages 1-2)
References
-
(hu2014kataninp80regulates pages 1-2): Wen F. Hu, Oz Pomp, Tawfeg Ben-Omran, Andrew Kodani, Katrin Henke, Ganeshwaran H. Mochida, Timothy W. Yu, Mollie B. Woodworth, Carine Bonnard, Grace Selva Raj, Thong Teck Tan, Hanan Hamamy, Amira Masri, Mohammad Shboul, Muna Al Saffar, Jennifer N. Partlow, Mohammed Al-Dosari, Anas Alazami, Mohammed Alowain, Fowzan S. Alkuraya, Jeremy F. Reiter, Matthew P. Harris, Bruno Reversade, and Christopher A. Walsh. Katanin p80 regulates human cortical development by limiting centriole and cilia number. Neuron, 84:1240-1257, Dec 2014. URL: https://doi.org/10.1016/j.neuron.2014.12.017, doi:10.1016/j.neuron.2014.12.017. This article has 124 citations and is from a highest quality peer-reviewed journal.
-
(peluso2021wholeexomesequencing pages 6-8): Francesca Peluso, Stefano Giuseppe Caraffi, Roberta Zuntini, Gabriele Trimarchi, Ivan Ivanovski, Lara Valeri, Veronica Barbieri, Maria Marinelli, Alessia Pancaldi, Nives Melli, Claudia Cesario, Emanuele Agolini, Elena Cellini, Francesca Clementina Radio, Antonella Crisafi, Manuela Napoli, Renzo Guerrini, Marco Tartaglia, Antonio Novelli, Giancarlo Gargano, Orsetta Zuffardi, and Livia Garavelli. Whole exome sequencing is the minimal technological approach in probands born to consanguineous couples. Genes, 12:962, Jun 2021. URL: https://doi.org/10.3390/genes12070962, doi:10.3390/genes12070962. This article has 4 citations.
-
(jin2017kataninp80numa pages 1-3): Mingyue Jin, Oz Pomp, Tomoyasu Shinoda, Shiori Toba, Takayuki Torisawa, Ken’ya Furuta, Kazuhiro Oiwa, Takuo Yasunaga, Daiju Kitagawa, Shigeru Matsumura, Takaki Miyata, Thong Teck Tan, Bruno Reversade, and Shinji Hirotsune. Katanin p80, numa and cytoplasmic dynein cooperate to control microtubule dynamics. Scientific Reports, Jan 2017. URL: https://doi.org/10.1038/srep39902, doi:10.1038/srep39902. This article has 47 citations and is from a peer-reviewed journal.
-
(hu2014kataninp80regulates pages 2-4): Wen F. Hu, Oz Pomp, Tawfeg Ben-Omran, Andrew Kodani, Katrin Henke, Ganeshwaran H. Mochida, Timothy W. Yu, Mollie B. Woodworth, Carine Bonnard, Grace Selva Raj, Thong Teck Tan, Hanan Hamamy, Amira Masri, Mohammad Shboul, Muna Al Saffar, Jennifer N. Partlow, Mohammed Al-Dosari, Anas Alazami, Mohammed Alowain, Fowzan S. Alkuraya, Jeremy F. Reiter, Matthew P. Harris, Bruno Reversade, and Christopher A. Walsh. Katanin p80 regulates human cortical development by limiting centriole and cilia number. Neuron, 84:1240-1257, Dec 2014. URL: https://doi.org/10.1016/j.neuron.2014.12.017, doi:10.1016/j.neuron.2014.12.017. This article has 124 citations and is from a highest quality peer-reviewed journal.
-
(brown2017geneticrequirementsfor pages 103-108): C Brown. Genetic requirements for building a brain of a sufficent size: insights from mendelian congenital microcephaly disorders. Unknown journal, 2017.
-
(peluso2021wholeexomesequencing pages 1-2): Francesca Peluso, Stefano Giuseppe Caraffi, Roberta Zuntini, Gabriele Trimarchi, Ivan Ivanovski, Lara Valeri, Veronica Barbieri, Maria Marinelli, Alessia Pancaldi, Nives Melli, Claudia Cesario, Emanuele Agolini, Elena Cellini, Francesca Clementina Radio, Antonella Crisafi, Manuela Napoli, Renzo Guerrini, Marco Tartaglia, Antonio Novelli, Giancarlo Gargano, Orsetta Zuffardi, and Livia Garavelli. Whole exome sequencing is the minimal technological approach in probands born to consanguineous couples. Genes, 12:962, Jun 2021. URL: https://doi.org/10.3390/genes12070962, doi:10.3390/genes12070962. This article has 4 citations.
-
(oegema2020internationalconsensusrecommendations pages 8-9): Renske Oegema, Tahsin Stefan Barakat, Martina Wilke, Katrien Stouffs, Dina Amrom, Eleonora Aronica, Nadia Bahi-Buisson, Valerio Conti, Andrew E. Fry, Tobias Geis, David Gomez Andres, Elena Parrini, Ivana Pogledic, Edith Said, Doriette Soler, Luis M. Valor, Maha S. Zaki, Ghayda Mirzaa, William B. Dobyns, Orly Reiner, Renzo Guerrini, Daniela T. Pilz, Ute Hehr, Richard J. Leventer, Anna C. Jansen, Grazia M. S. Mancini, and Nataliya Di Donato. International consensus recommendations on the diagnostic work-up for malformations of cortical development. Nature Reviews Neurology, 16:618-635, Sep 2020. URL: https://doi.org/10.1038/s41582-020-0395-6, doi:10.1038/s41582-020-0395-6. This article has 130 citations and is from a highest quality peer-reviewed journal.
-
(oegema2020internationalconsensusrecommendations pages 7-8): Renske Oegema, Tahsin Stefan Barakat, Martina Wilke, Katrien Stouffs, Dina Amrom, Eleonora Aronica, Nadia Bahi-Buisson, Valerio Conti, Andrew E. Fry, Tobias Geis, David Gomez Andres, Elena Parrini, Ivana Pogledic, Edith Said, Doriette Soler, Luis M. Valor, Maha S. Zaki, Ghayda Mirzaa, William B. Dobyns, Orly Reiner, Renzo Guerrini, Daniela T. Pilz, Ute Hehr, Richard J. Leventer, Anna C. Jansen, Grazia M. S. Mancini, and Nataliya Di Donato. International consensus recommendations on the diagnostic work-up for malformations of cortical development. Nature Reviews Neurology, 16:618-635, Sep 2020. URL: https://doi.org/10.1038/s41582-020-0395-6, doi:10.1038/s41582-020-0395-6. This article has 130 citations and is from a highest quality peer-reviewed journal.
-
(hu2014kataninp80regulates media 9e221719): Wen F. Hu, Oz Pomp, Tawfeg Ben-Omran, Andrew Kodani, Katrin Henke, Ganeshwaran H. Mochida, Timothy W. Yu, Mollie B. Woodworth, Carine Bonnard, Grace Selva Raj, Thong Teck Tan, Hanan Hamamy, Amira Masri, Mohammad Shboul, Muna Al Saffar, Jennifer N. Partlow, Mohammed Al-Dosari, Anas Alazami, Mohammed Alowain, Fowzan S. Alkuraya, Jeremy F. Reiter, Matthew P. Harris, Bruno Reversade, and Christopher A. Walsh. Katanin p80 regulates human cortical development by limiting centriole and cilia number. Neuron, 84:1240-1257, Dec 2014. URL: https://doi.org/10.1016/j.neuron.2014.12.017, doi:10.1016/j.neuron.2014.12.017. This article has 124 citations and is from a highest quality peer-reviewed journal.
-
(lynn2021themammalianfamily pages 1-2): Nicole A. Lynn, Emily Martinez, Hieu Nguyen, and Jorge Z. Torres. The mammalian family of katanin microtubule-severing enzymes. Frontiers in Cell and Developmental Biology, Aug 2021. URL: https://doi.org/10.3389/fcell.2021.692040, doi:10.3389/fcell.2021.692040. This article has 50 citations.
-
(lynn2023methodsandsystems pages 20-26): NA Lynn. Methods and systems developed for characterization of mammalian katanin. Unknown journal, 2023.
-
(peluso2021wholeexomesequencing pages 4-6): Francesca Peluso, Stefano Giuseppe Caraffi, Roberta Zuntini, Gabriele Trimarchi, Ivan Ivanovski, Lara Valeri, Veronica Barbieri, Maria Marinelli, Alessia Pancaldi, Nives Melli, Claudia Cesario, Emanuele Agolini, Elena Cellini, Francesca Clementina Radio, Antonella Crisafi, Manuela Napoli, Renzo Guerrini, Marco Tartaglia, Antonio Novelli, Giancarlo Gargano, Orsetta Zuffardi, and Livia Garavelli. Whole exome sequencing is the minimal technological approach in probands born to consanguineous couples. Genes, 12:962, Jun 2021. URL: https://doi.org/10.3390/genes12070962, doi:10.3390/genes12070962. This article has 4 citations.
-
(lynn2023methodsandsystemsa pages 31-36): NA Lynn. Methods and systems developed for characterization of mammalian katanin. Unknown journal, 2023.
-
(lynn2023methodsandsystems pages 41-46): NA Lynn. Methods and systems developed for characterization of mammalian katanin. Unknown journal, 2023.
-
(zaidi2022primaryciliainfluence pages 9-10): Donia Zaidi, Kaviya Chinnappa, and Fiona Francis. Primary cilia influence progenitor function during cortical development. Cells, 11:2895, Sep 2022. URL: https://doi.org/10.3390/cells11182895, doi:10.3390/cells11182895. This article has 28 citations.
-
(duy2024the"microcephalichydrocephalus" pages 6-7): Phan Q. Duy, Neel H. Mehta, and Kristopher T. Kahle. The "microcephalic hydrocephalus" paradox as a paradigm of altered neural stem cell biology. Cerebral cortex, Nov 2024. URL: https://doi.org/10.1093/cercor/bhad432, doi:10.1093/cercor/bhad432. This article has 4 citations and is from a highest quality peer-reviewed journal.
-
(oegema2020internationalconsensusrecommendations pages 12-13): Renske Oegema, Tahsin Stefan Barakat, Martina Wilke, Katrien Stouffs, Dina Amrom, Eleonora Aronica, Nadia Bahi-Buisson, Valerio Conti, Andrew E. Fry, Tobias Geis, David Gomez Andres, Elena Parrini, Ivana Pogledic, Edith Said, Doriette Soler, Luis M. Valor, Maha S. Zaki, Ghayda Mirzaa, William B. Dobyns, Orly Reiner, Renzo Guerrini, Daniela T. Pilz, Ute Hehr, Richard J. Leventer, Anna C. Jansen, Grazia M. S. Mancini, and Nataliya Di Donato. International consensus recommendations on the diagnostic work-up for malformations of cortical development. Nature Reviews Neurology, 16:618-635, Sep 2020. URL: https://doi.org/10.1038/s41582-020-0395-6, doi:10.1038/s41582-020-0395-6. This article has 130 citations and is from a highest quality peer-reviewed journal.
-
(lynn2021themammalianfamily pages 14-15): Nicole A. Lynn, Emily Martinez, Hieu Nguyen, and Jorge Z. Torres. The mammalian family of katanin microtubule-severing enzymes. Frontiers in Cell and Developmental Biology, Aug 2021. URL: https://doi.org/10.3389/fcell.2021.692040, doi:10.3389/fcell.2021.692040. This article has 50 citations.
-
(lynn2021themammalianfamily pages 8-10): Nicole A. Lynn, Emily Martinez, Hieu Nguyen, and Jorge Z. Torres. The mammalian family of katanin microtubule-severing enzymes. Frontiers in Cell and Developmental Biology, Aug 2021. URL: https://doi.org/10.3389/fcell.2021.692040, doi:10.3389/fcell.2021.692040. This article has 50 citations.
-
(lynn2023methodsandsystemsa pages 41-46): NA Lynn. Methods and systems developed for characterization of mammalian katanin. Unknown journal, 2023.
-
(duy2024the"microcephalichydrocephalus" pages 4-5): Phan Q. Duy, Neel H. Mehta, and Kristopher T. Kahle. The "microcephalic hydrocephalus" paradox as a paradigm of altered neural stem cell biology. Cerebral cortex, Nov 2024. URL: https://doi.org/10.1093/cercor/bhad432, doi:10.1093/cercor/bhad432. This article has 4 citations and is from a highest quality peer-reviewed journal.
-
(oegema2020internationalconsensusrecommendations pages 1-2): Renske Oegema, Tahsin Stefan Barakat, Martina Wilke, Katrien Stouffs, Dina Amrom, Eleonora Aronica, Nadia Bahi-Buisson, Valerio Conti, Andrew E. Fry, Tobias Geis, David Gomez Andres, Elena Parrini, Ivana Pogledic, Edith Said, Doriette Soler, Luis M. Valor, Maha S. Zaki, Ghayda Mirzaa, William B. Dobyns, Orly Reiner, Renzo Guerrini, Daniela T. Pilz, Ute Hehr, Richard J. Leventer, Anna C. Jansen, Grazia M. S. Mancini, and Nataliya Di Donato. International consensus recommendations on the diagnostic work-up for malformations of cortical development. Nature Reviews Neurology, 16:618-635, Sep 2020. URL: https://doi.org/10.1038/s41582-020-0395-6, doi:10.1038/s41582-020-0395-6. This article has 130 citations and is from a highest quality peer-reviewed journal.
-
(duy2024the"microcephalichydrocephalus" pages 1-2): Phan Q. Duy, Neel H. Mehta, and Kristopher T. Kahle. The "microcephalic hydrocephalus" paradox as a paradigm of altered neural stem cell biology. Cerebral cortex, Nov 2024. URL: https://doi.org/10.1093/cercor/bhad432, doi:10.1093/cercor/bhad432. This article has 4 citations and is from a highest quality peer-reviewed journal.